頁概要 (Pdfファイル)
◆頁1 《ヒメオニササガヤ詳細図》
◆頁2 《用語・略語》 《解説》
◆頁3 《ノート》 《文献》
特徴的な部位の部分図 (2004東京都府中市・2010東京都国立市 採集品) (Pdfファイル→ヒメオニササガヤ1詳細図 頁1)
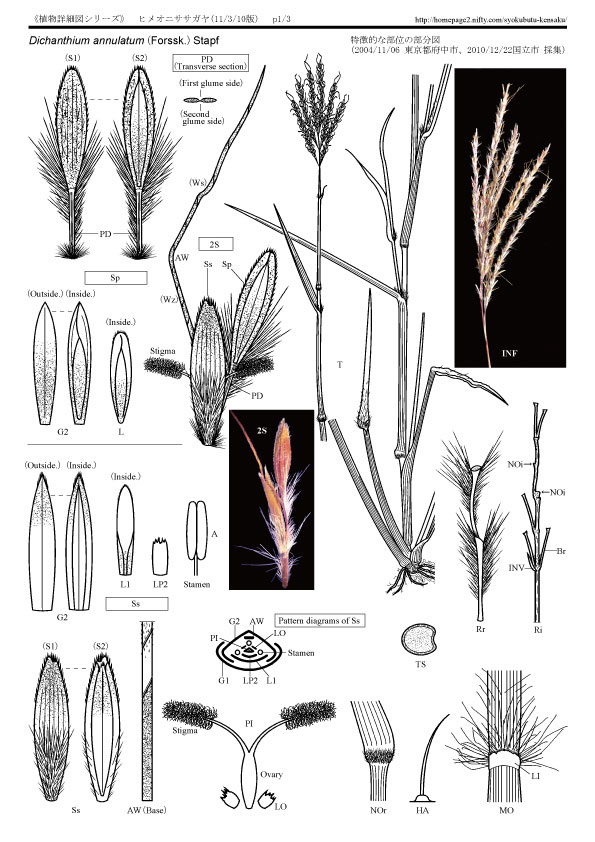 《形態学用語その他の略語は長田に順じた》(ABC順)
《形態学用語その他の略語は長田に順じた》(ABC順)A:Anther(葯)
AW:Awn(芒)
:Base(基部)
Br:Branch(A raceme was removed.)(花序の枝:総を取外した)
G1:First glume(第一苞穎)
G2:Second glume(第二苞穎)
HA:Hair(Papilla-based)(毛:基部が乳頭状の)
INF:Inflorescence(花序)
Inside.(内側)
INV:Rudimentary lnvolucre(退化して痕跡的になった総苞)
L:Lemma(護穎)
L1:Lemma of first floret(第一小花の護穎)
LI:Ligule(葉舌)
LO:Lodicule(りん皮)
LP2:Palea of second floret(第二小花の内穎)
MO:Mouth of leaf-sheath(葉鞘口部)
NOc:Node of culm(稈の節)
NOi:Node of inflorescence(花序の節)
Outside.(外側)
Ovary(子房)
Pattern diagrams of Ss(無柄小穂の模式図)
PD:Pedicel(小穂の柄)
:First glume side(第一苞穎側)
:Second glume side(第二苞頴側)
:Transverse section(横断面)
PI:Pistil(雌しべ)
Ri:Rachis of inflorescence(花序の中軸)
Rr:Rachis of raceme(A Sp and a Ss were removed.)(総の中軸:有柄小穂と無柄小穂を取り除いた)
2S:Paired spikelets(有柄と無柄の対になった小穂)
S1:Spikelet seen from first glume side(第一苞穎側からみた小穂)
S2:Spikelet seen from second glume side(第二苞穎側からみた小穂)
Sp:Pedicellate spikelet(有柄の小穂)
Ss:Sessile spikelet(無柄の小穂)
Stamen(雄しべ)
Stigma(柱頭)
T:Plant(most1y total view)(植物体)
TS:Transverse section of culm(稈の断面)
Ws:The S type is wind(S巻、S型巻き)
Wz:The Z type is wind(Z巻、Z型巻き)
この頁top
ヒメオニササガヤ Dichanthium annulatum (Forssk.) Stapf. (2011/3/10) (Pdfファイル→ヒメオニササガヤ解説 頁2 頁3)
本種は、小穂は2小花で構成され上方の第2小花が稔性を持つ・苞穎が護穎より発達する・花序中軸の各節には形状の異なる2小穂が対となる、等の特徴でキビ亜科ヒメアブラススキ連に含まれるイネ科植物である。オニササガヤ属 Dichanthium は本検索サイトでの属タイプ「1E型」、【構成2小花;1小花+1頴+苞穎(小穂と同長)】で、小穂略記は[1小花+1頴+長苞]。本種は現時点で東京都・兵庫県・沖縄県で記録されている。長田(1993)に本種の掲載はない。
《解説》 (2004/11/6東京都府中市・2010/12/22東京都国立市 採集品による)
◇多年草。少数の稈が叢生してやや少群生する。高さ7~80(100)cm、稈は中実、節には斜上する短毛が密生する。
◇葉身は長いもので18cm、幅は2~4mm。葉には基部の膨れた毛があり、葉表や基部に多い。
◇葉鞘は平滑、無毛。葉舌は膜状で高さ1~1.5mm、上縁は短毛列状。
◇花序は稈に頂生し長さ5~7cm。枝先や中軸の節に斜上直立した総をつけ、やや掌状の花序となる。花序ははじめ淡緑色ですぐ紫褐色となり、熟期には白味を帯びる。
・花序の中軸は長さ3~4cm(注)、中軸に圧閉して斜上直立する短い枝を2~5個ほどつける。中軸と枝はともに紫褐色でほぼ無毛だが、枝腋に毛叢がある。
・総は小枝の先と花序中軸の節につき、2~8個が斜上直立する。
・総の中軸の各節の左右には、無柄と有柄の小穂が対となって互生するが、先端部のみ無柄小穂1個と有柄小穂が2個の組み合わせとなる。節には関節があり、無柄小穂・有柄小穂・ひと節分の総の中軸がセットとなり熟期に脱離する。
・総の中軸はやや扁平で節毎に少し湾曲し、両側縁には斜上する半透明の白長毛が1列に列生する。節の関節から上の部分が脱落すると、先端は杯状のくぼみとなる。
(注)桑原(2008)には円錐花序で中軸が6~15cmと長く、葉の長さ15~30cm、幅4~8mmと全体大振りな個体の図があり、本土のものとは少し外観が異なるタイプである(東京都産ではやや掌状の総状花序で、花序中軸は長さ3~4cm、葉の長さ18cm、葉幅2~4mm)。
◇無柄小穂は2小花からなり、第一小花は不稔でほぼ無性。第二小花は稔性を持ち両性で屈曲した長芒を持つ。
・第一苞穎は小穂と同大同長で長さ約4mm、扁平な細楕円形で鈍円頭、紫渇色で数脈があり下半部に半透明の白直毛が多数ある。左右の縁は内折れし2竜骨状、上部の竜骨には刺毛が密に並ぶ。
・第二苞穎は小穂とやや同長。芒の位置する背側が突出し中央に緑色の1脈があり鋭頭、光沢のある膜質で上部縁の周辺に軟毛がある。両縁は内折れしやや2竜骨状。
・第一小花は護穎のみに退化し半透明の薄膜質で無脈、長さ約2.5~3mmで下半部はやや紫色を帯び、下方の両縁はゆるく内折する。
・第二小花は両性で護穎は膜質部がなく基部は上部と同幅で少し厚みが薄く、全体が芒状で長さは16mm前後。強く捩れ中央部でくの字状に屈曲する。第二小花の護穎は形状不定な微小薄膜質または消失。
◇有柄小穂は無芒で不稔。第一苞穎は長さ約3.8mmで形や色は無柄小穂のものと同様であるが下半部の毛は少ない。やや小さく、乾燥すると無柄小穂より幅は狭くなる。第二苞穎は膜質で長さ約3.3mm、左右の縁が内折れし下半部は紫色を帯びる。護穎は長さ約2.5mmで左右の縁が内折れし下半部は紫色を帯びる。
・有柄小穂の柄は両側縁に斜上する半透明の白直毛が1列に列生し、毛は柄の上部ほど長くなる。柄には縦中央に溝があり、溝部分は薄質でやや半透明状となり、溝は第一苞穎側がより深い。
◇雌しべ1個、花柱2個、柱頭は2個で濃紫色のブラシ状。
◇雄しべ3個。花糸は底着する。
◇鱗被は薄膜質で2個。 この頁top
《ノート》
木場(2003)・植村 他(2010)によると、沖縄県に牧草として導入されたものが逸出し、兵庫県で1995年頃採集され和名が新称された。オニササガヤ属 Dichanthium は日本に自生がなく、本種のほかにオニササガヤ D. aristatum (Poir.) C.E.Hubb. 、シラゲオニササガヤ D. sericeum (R.Br.) A.Camus が知られ、いずれも帰化植物である。
◇本種の芒は第二小花の護穎の変化したものであるが、基部の頴にあたる部分が完全に退化して全体が1本の芒状となり、大変特徴的である。このような芒を持つものは他には在来種であるヒメアブラススキ Capillipedium parviflora (R.Br.) Stapf 、メガルカヤ Themeda triandra Forssk. var. japonica (Willd.) Makino などが知られる。
◇芒は基部を除く下半部でZ巻、上半部がS巻に捩れるため、中央部でくの字状に屈曲する。芒の上下で捩れ方向が異なる例はイネ科植物ではよくみられる。屈曲する芒の役割は散布後に、湿乾による芒の回転運動により種子を地中に埋め込む作用を持つと考えられている。
◇有柄小穂の柄の半透明状の縦溝は、大変特異な形質である(図 PD:Transverse section)。
杉本(1973)はヒメアブラススキ属の特徴として、「花軸と小梗(小穂の柄)に透明条または溝条がある」とし、木村(2003)はヒメアブラススキ属・モンツキガヤ属 Bothriochloa の特徴として、「小穂の柄と節間は半透明の中央線をもつ」としている。
小穂の柄の半透明状の縦溝がオニササガヤ属の特徴であると仮定して、主要な文献のイネ科の検索表の中でのオニササガヤ属の位置を検証してみると以下のようになる。小山(1964)・杉本(1973)・木村(2003)は、小花の形質→小穂の形質→花序の形質の順に検索が行われ、オニササガヤ属もこれら属の近くに位置する(木場2001は、総苞葉の形質→花序の形質と検索が進むため、オニササガヤ属はこの段階で別れるため、ここでは検証を省く)。小穂の柄の半透明状の縦溝は柄を透かすと確認できるが、どのような役割を持つものなのかは筆者にはまだ理解できていない。
・杉本(1973)
◆G花軸と小梗に透明条又は溝条がある『ヒメアブラススキ属』(モンツキガヤ属を分けていないため、モンツキガヤのみ考慮した)
◆b花序の枝は更に2~3回複分岐する;小枝は開出、総は1~8関節あり、各総は大部分細い、2~8mmの小梗がある【ヒメアブラススキ】
◆b花序の枝は単純なるか1~8小枝を出す;小枝は上向して鋭角又は閉伏する;総に短柄があり、5~多関節【モンツキガヤ】
(ヒメオニササガヤの総には柄がないと思うので、この点で区別できると考える)
・木村(2003)
◆E花序は1つの総かやや掌状;時に伸張した花軸をもつが、その場合節間は半透明の中央線をもつ
◆F小穂の柄と節間は半透明の中央線をもつ◆G円錐花序『ヒメアブラススキ属』
◆G花序は単一かやや掌状『モンツキガヤ属』(ここに位置すると考えられる)
・小山(1964)
◆3.登実小穂の第1苞穎は草質
◆4.葉狭く長い◆5.総のすべての対小穂は皆異形;第1小穂は雄性;第2小穂は両性、稀に雌性
◆6.花序は稈の頂だけにつく、その基部に花序を抱く苞がない『ヒメアブラススキ属』
(ヒメオニササガヤはやや掌状花序で、円錐花序のヒメアブラススキと区別できる)
◇イネ科植物は中空の稈をもつものが一般的だが、本種の稈は中実である。ジュズダマ属(Coix)やワセオバナ属(Saccharum)など、中実の稈をもつものもいくつか知られ、本種を含め比較的熱帯原産の種が多いように思われる。
◇本種の花序の基部には、ごくまれに小片状のものがみられる(図 INV)。いくつかのイネ科植物では、花序の基部に切込状の括れや小片状のものが痕跡として残っているのがみられ、進化の過程で存在していた総苞が退化したものとも考えられ、本種もその一つであろう。イネ科でも総苞を持つ種類がいくつかあるが、いずれも花序を保護する機能に特化したもので、イグサ科やカヤツリグサ科の植物にみるような、葉身の発達した総苞葉と呼ばれるものではない。
◇果実が熟す頃は小穂の紫渇色が脱色して白くなり、総の中軸の節の関節で分離する小穂のセットが、風に吹かれて飛び散り種子散布される。小柄などの半透明の長い白長毛は、風に乗せて遠くへ運ばせる作用を多少負っていると考えられる。
◇日本に帰化しているオニササガヤ属の種について記しておくと、
・オニササガヤ:植村 他(2010)によると沖縄県に帰化しており、穂状花序の柄が有毛であるという。
・シラゲオニササガヤ:木場(2003)によると愛知県で1959年に記録され、走出枝があり、葉舌の長さ1.5mm未満と短いことでヒメオニササガヤと区別できるという。植村 他(2010)によると三重県にも記録があり、第一苞穎の上部に長毛が密生するという。(2011/3/10:山口純一)
《参考文献》
木村陽子 2003.イネ科.千葉県の自然誌 別編4 千葉県植物誌,pp.711-787.千葉県.
木場英久 2001.イネ科検索表.神奈川県植物誌2001,pp.257-262.神奈川県立生命の星・地球博物館.
木場英久 2003.イネ科オニササガヤ属.日本の帰化植物,pp.288-290.平凡社.
小山鐡夫 1964.イネ科.原色日本植物図鑑 草本編Ⅲ,pp.301-388.保育社.
桑原義晴 2008.桑原義晴 日本イネ科植物図譜,503pp.全国農村教育協会.
杉本順一 1973.日本草本植物総検索誌Ⅱ単子葉編,630pp.井上書店.
植村修二他 2010.日本帰化植物写真図鑑 第2巻,576pp.全国農村教育協会.
この頁top
syokubutu kensaku