《植物詳細図シリーズ》 スズメノナギナタ Parapholis incurva (L.)C.E.Hubbard 植物詳細図top HPtop イネ科top zusyousai003
特徴的な部位の部分図 (採:07/6/6 東京都) 解説 進化想像図
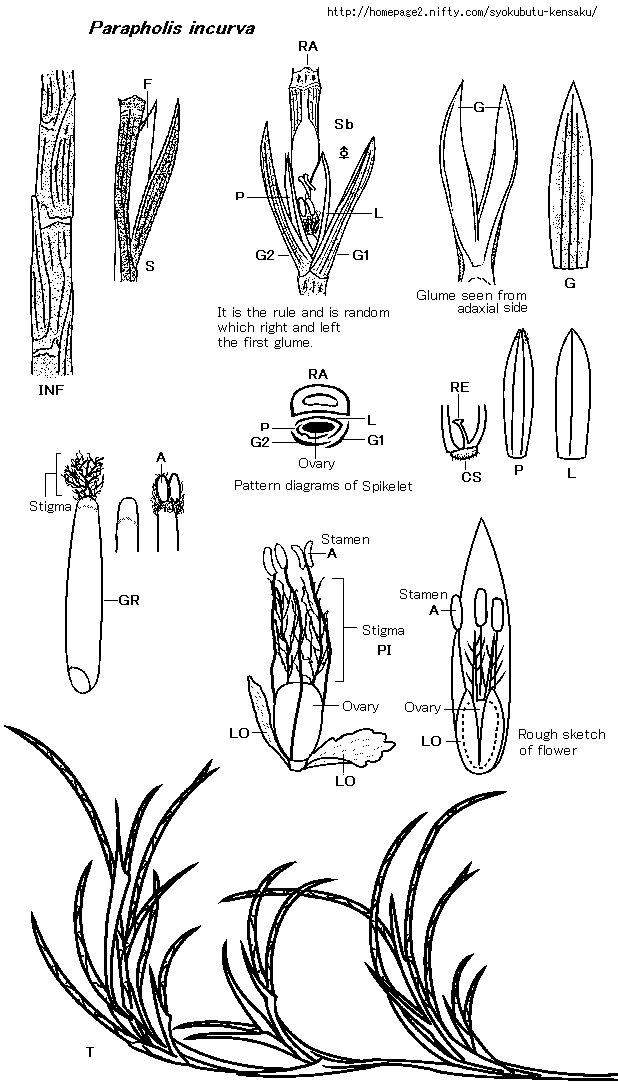 《形態学用語その他の略語は長田1993に順じた》(ABC順)
《形態学用語その他の略語は長田1993に順じた》(ABC順)
A: Anther(葯)
CS: Callus(小花の基盤)
F: Floret(小花)
G: Glume(苞穎)
:Glume seen from adaxial side(花軸側(向軸面)からみた苞穎)
GR: Grain(caryopsis)(果実;穎果)
INF: A part of inflorescence(花序の一部)
It is the rule and is random which right and left the first glume.(左右どちらが第一苞頴かは規則性がなくランダム)
L: Lemma(護穎)
LO: Lodicule(りん皮)
Ovary(子房)
P: Palea(内穎)
Pattern diagrams of Spikelet(小穂の模式図)
PI: Pistil(雌しべ)
RA: Rachis of inflorescence(花序の中軸)
RE: Rachilla extention(小軸突起)
Rough sketch of flower(花の略図)
S: Spikelet(小穂)
Sb: Spikelet seen from abaxial side(花軸と反対側(背軸面)からみた小穂)
Stamen(雄しべ)
Stigma(柱頭)
T: Plant(mostly total view (植物体;大部分は全形図)
この頁top
スズメノナギナタ Parapholis incurva (L.) C.E.Hubbard 図 進化想像図
本種はイネ科スズメノナギナタ属で、鎌状に曲がった花序の外観から、同定は容易である。
総は稈頂に1本で花序中軸は太く小穂と同巾以上で半ば小穂が埋まり、本検索サイトでの属タイプ「7C型・7D型」となる。
【構成1小花;1小花+苞穎(小穂と同長)】で、小穂略記は[1小花+長苞]である。
《各部分の形態》
◇植物体は基部からすぐに苞葉状の葉鞘中から数本枝を分岐し、枝は短くすぐ上の葉鞘中から更に数本枝分かれを繰り返し、総じて基部から著しい数の枝を分岐する。各枝は短く、葉鞘の中から長い総状花序を1本頂部につける。総状花序は半円を描くように鎌曲し特異である(図T)。
◇小穂は花序中軸に半ば埋まるように互生し、総は円筒状を呈し径1.5mm(図INF)、その中軸部は中空で特異である(図RA)。
◇苞頴は小穂と同長で2個。中軸に対し並列していて(図G、G1、G2)、これだけで属が決まるほど特異である。長さ5〜6mm、硬草質で、合わせ目側は白膜縁、上部外側縁に微刺が疎らにある。背に3〜4本の縦脈があり、花軸と同様に縦脈の隆起が目立つ。第一苞頴と第二苞頴は合わせ目で少し重なり、第一苞頴が外側にある。
◇小穂は1小花で、護穎はやや硬い膜質、長さ3.8〜4.5mm、1脈で上部はやや竜骨気味(図L)。内頴は極薄い膜質で、上部の背に微毛があり、長さ3.2〜4mm、、先端は浅く2裂し、背に2脈ありその上部はやや竜骨気味(図P)。
◇極薄膜質の鱗被が2個あり、長さ0.8〜1.1mmで小花の1/4ほどもあり、他種に比べてかなり大きく目立つ(図LO)(鱗被については、トピック「イネ科各種の受粉の秘密」に解説あり)。本種の鱗被は、子房が若い時点では、子房や雄しべ・雌しべをスッポリと抱きかかえるように包んでいるが、やがて雄しべ・雌しべが上部隙間から鱗被の外に伸びだす(図Rough sketch of flower)。小花が薄膜質で有る事などから、開花の役としては働いていないかも知れない。小花の開花にあたっては硬い苞頴がまず左右に斜開するが、鱗被が作用しているとは思われず、植物体の乾燥による作用が働いているのではないかと想像するが、定かではない。
◇小花の基盤は0.2mm程で有毛である(図CS)。小軸突起がある(図RE)。◇頴果の長さ3.2mm(図GR)。
◇雄しべ3個。葯は黄色で、長さ0.4〜0.9mm(図A)。花糸は極細い毛状で長さ2.5mm。葯の形態は次の状態が見られた。
1.苞頴が閉じていて、小穂の外に葯が出ているもの。他家受粉を目的としている。
2.花糸が伸びきらず、若い子房の頂部に柱頭と共に接したままのもの、多く見られるがこれが閉鎖花に該当するものと考える。
3.子房が若い時点ですでに花糸が伸びきっているもの(図Stamen)、子房が成長してくると柱頭に葯が接する。
頴果の頂部に毛玉と成った柱頭と葯が接したままのものが目に付くが、2.3.のタイプが最終的にこうなる(図GRの右端)。
―――――――――――――――――――――――――――――――――――
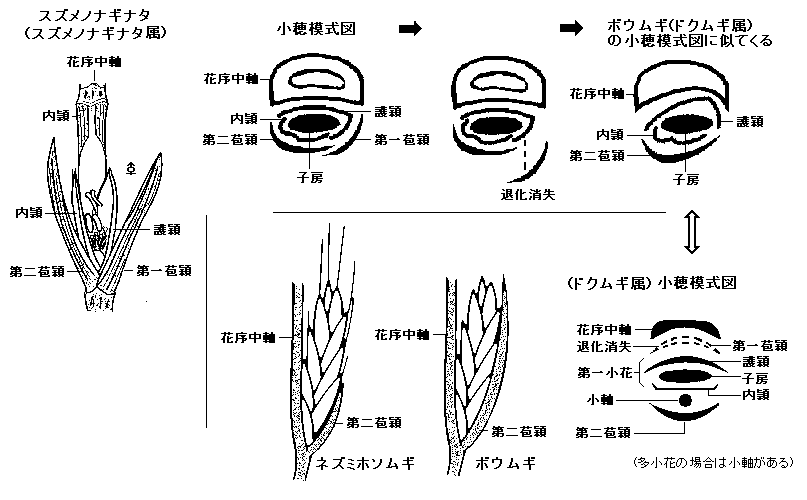
◆本種において形態的に最も注目したいのは、小穂各部分の微妙な位置関係についてである。
◇まず疑問におもったのは、並列している2枚の苞頴の、左右どちらが第一苞頴で、どちらが第二苞頴であるのかについて調べてみたが、通常なら規則的に成っていると考えるのが自然なはずだが、本種においては全く不規則であることが判明した(図t is the rule and is random which right and left the first glume.)。
次に護穎と内頴の位置と、苞頴との関連が正しく保たれているかを調べてみた。その結果、護穎は必ず第一苞頴の側に背を向けていて、内頴は第二苞頴側に背を向けていた。これは理屈どおりであり問題はなかった。
しかし、(図Pattern diagrams of Spikelet)を良くみていただきたい。第一苞頴が右にあろうと左にあろうと、護穎は必ず花軸側に位置するのである。そして内頴は外側に配置している。
これを形態的にのみ思考した場合、本種は何れ第一苞頴は不要と成り消失すると仮定すれば、第二苞穎が花軸の前面に位置してドクムギ属の小穂形態と同じに成る。特に外観もボウムギと非常に良く似た形と成りうる(下の、「ボウムギの構造に似てくる進化想像図」参照)。いいかえれば、第一苞頴が消失して、その役割を花軸側が負う構造に進化した形のドクムギ属の、一歩手前の状態を暗示させるような、非対称に歪んだ模式図を持っているのである。
何れにしろ苞頴が花軸の前面に並列的に並ぶ姿は極めて特異である。
もうひとつ。筆者が本種を採集したのは07/6/6日で、丁度開花の時期であった。それから2週間後の6/20日におとづれると、総ての個体がドライフラワー状態で枯れていて、触ると細かく砕けてしまうのである。勿論周囲の植物に異変は見られない。原因は乾燥だと思うのだが、生育場所的なものなのか、或いはこのような生態的特長を持っている植物なのかは判然としないが、少し早過ぎる気がするが、この点は全く理解できない。(07/6/28 山口純一)
《主な参考文献》
木村陽子 2003.イネ科.千葉県の自然誌 別編4 千葉県植物誌,pp.711-787.千葉県.
木場英久 2001.イネ科 スズメノナギナタ属.神奈川県植物誌2001,pp.278-279.神奈川県立生命の星・地球博物館.
木場英久 2003.イネ科 スズメノナギナタ属.日本の帰化植物,p.252.平凡社.
長田武正 1993.増補 日本イネ科植物図譜,776pp.平凡社.
杉本順一 1973.日本草本植物総検索誌Ⅱ単子葉編,630pp.井上書店. スズメノナギナタtop 進化想像図
(ボウムギの構造に似てくる進化想像図)
 スズメノナギナタtop 解説
スズメノナギナタtop 解説
syokubutu kensaku